Producción secundaria
La producción secundaria es un concepto de ecosistema (flujo de energía) que se mide utilizando una técnica de población. Los libros de texto rara vez mencionan que también pueden abordar cuestiones de población / comunidad.

Introducción
La producción secundaria representa la formación de masa viva de una población o grupo de poblaciones heterótrofas durante un período de tiempo (Benke y Huryn 2006). Es el equivalente heterótrofo de la producción primaria neta por autótrofos. Llevada al extremo, la producción secundaria puede representar la formación de masa para todo un nivel trófico. La producción animal, el tema de este artículo, casi siempre se mide a nivel de población, independientemente de si se considera una sola población, un grupo de poblaciones o un nivel trófico completo. Si se busca medir la producción de un nivel trófico completo, se debe sumar la producción de todas las poblaciones dentro de ese nivel, o al menos las principales. A diferencia de,
La producción secundaria históricamente se ha visto en el contexto del flujo de energía a través de los niveles tróficos (p. Ej., Odum 1971). Los primeros estudios de flujo de energía utilizaron medidas energéticas (kilocalorías o kilojulios). Sin embargo, la mayoría de las estimaciones de la producción actual, ya sea para productores primarios (autótrofos) o productores secundarios (heterótrofos), se expresan como masa (gramos de carbono o gramos de masa seca). Si bien las unidades de biomasa de la población a menudo se presentan como gramos / m 2 , la unidad típica para la producción secundaria incorpora el tiempo (por ejemplo, gramos m -2 año -1 , gramos m -2 semana -1). Tendemos a pensar en la biomasa como una variable estructural (o estática) y la producción como una variable funcional porque esta última mide un proceso ecológico a través del tiempo.
Se ha reconocido desde hace mucho tiempo que no todos los alimentos ingeridos por un individuo se convierten en nueva masa animal. Considere un caracol de arroyo pastando en algas (Figura 1). Sólo una fracción del material ingerido (I) se asimila (A) del tracto digestivo; el resto se desmaya como heces (F). Del material asimilado, solo una fracción contribuye al crecimiento de la masa de un individuo o a la reproducción, los cuales, en última instancia, representan la producción (P). La mayor parte del resto se usa para la respiración (R). Una pequeña porción de energía se pierde en la excreción, pero generalmente se ignora en tales presupuestos de energía. Se utilizan ecuaciones simples para ilustrar el destino de la energía ingerida, como I = R + P + F. Alternativamente, la producción es P = I – F – R.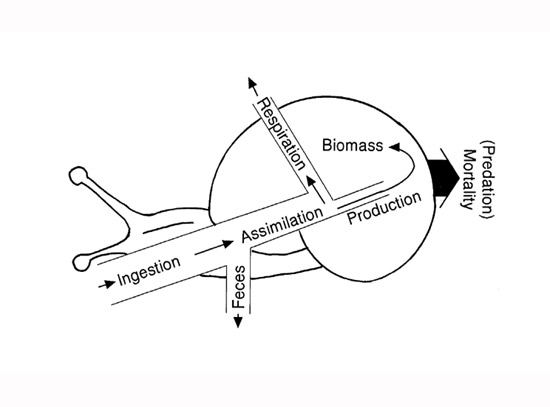
Figura 1: Diagrama de flujo de energía de una población de caracoles de arroyoTodos los flujos (ingestión, asimilación, heces, respiración, producción, mortalidad) tienen unidades de mg de masa seca m -2 día -1 ). El área ocupada por el caracol representa la biomasa (mg de masa seca m -2 ). La producción se suma a la biomasa y la mortalidad se resta de la biomasa simultáneamente. Por lo tanto, si la producción supera la biomasa perdida por mortalidad, la biomasa aumentará. Si la producción es menor que la biomasa perdida por mortalidad, la biomasa disminuirá.
La mayoría de los diagramas de flujo de energía en los libros de texto muestran que la producción fluye de un nivel trófico a otro. En realidad, sin embargo, la producción es un proceso de crecimiento que primero debe agregarse a la masa de individuos antes de que pueda consumirse en el proceso de fluir hacia un depredador o el siguiente nivel trófico (Figura 1). Por lo tanto, es un proceso de dos pasos: el crecimiento individual primero crea una nueva biomasa de población, luego parte de la biomasa fluye al siguiente nivel como una porción de los individuos consumidos por los depredadores.
La apreciación generalizada de la producción secundaria surgió en la década de 1960 con el establecimiento del Programa Biológico Internacional (IBP). Un enfoque principal del IBP fue cuantificar el flujo de energía a través de los niveles tróficos en varios ecosistemas, tanto acuáticos como terrestres, en todo el mundo. Los manuales del IBP proporcionaron los métodos básicos para estimar la producción (por ejemplo, Edmondson y Winberg 1971). No fue hasta los años setenta y ochenta que los ecologistas comenzaron a ajustar los métodos y abordar preguntas fuera del paradigma del flujo de energía (Benke y Huryn 2010). Es decir, la producción de población por sí sola (sin respiración, asimilación, etc.) pasó a utilizarse como variable de respuesta.
Un método simple para medir la producción secundaria
Este enfoque simple tiene en cuenta las pérdidas de biomasa debido a la mortalidad, así como la acumulación de biomasa a través del crecimiento individual. En este ejemplo particular, como ocurriría con un insecto, la producción somática solo ocurre durante las etapas juveniles y ninguna durante la etapa adulta. Los adultos producen huevos, pero esta contribución a la producción es a menudo pequeña (dependiendo del taxón) y se mediría con un método diferente, si es que se mide. Los cálculos de la producción real se realizan fácilmente en una tabla u hoja de cálculo como se muestra para los estadios larvarios de la libélula Epitheca spp. de un estanque (Tabla 1). Se suma la producción de todos los intervalos de tiempo para obtener la producción anual (1927,6 mg m -2 y -1). Este enfoque de estimación de la producción de cohortes se puede aplicar a otros tipos de historias de vida, incluso a aquellas en las que los juveniles crecen dentro de una bolsa de cría o útero.

Cuadro 1: Cálculo de la producción anual (o cohorte) de la libélula Epitheca spp (dos especies indistinguibles combinadas) de un estanque de Carolina del SurLa producción es la suma de la columna ÑΔW (modificada de Benke 1976).
La producción no es solo para el flujo de energía
Aunque la mayoría de los textos sobre ecología discuten la producción secundaria solo en la medida en que se aplica a los estudios de flujo de energía de los ecosistemas, muchas aplicaciones en las últimas décadas han tenido diferentes propósitos, particularmente para invertebrados bentónicos de agua dulce (Benke & Huryn 2010). Aquí se describen brevemente ejemplos de aplicaciones que abordan cuestiones energéticas y ecológicas.
- ¿Cómo se puede utilizar la producción con eficiencias ecológicas para comprender mejor el flujo de energía? Las eficiencias ecológicas básicas son la eficiencia de asimilación (asimilación / ingestión o A / I), la eficiencia de producción neta (producción / asimilación o P / A) y la eficiencia de producción bruta (producción / ingestión o P / I). Los cálculos de I, A, R y P generalmente se realizan en el laboratorio utilizando varios métodos. El crecimiento de la masa individual en el laboratorio se utiliza como sustituto de la producción en el campo. Podemos calcular que si se encuentra que un herbívoro (como un caracol, Figura 1) tiene una eficiencia de asimilación del 25% y una eficiencia de producción neta del 40%, entonces su eficiencia de producción bruta (A / I x P / A) es solo 0.25 x 0.40 = 0.10, o 10%. Así, si toda la materia orgánica producida por las algas fuera consumida por los caracoles, La producción de caracoles solo podría representar el 10% de la producción de redes de algas. Una lección obvia aprendida es que para cualquier parcela de tierra o volumen de agua, solo una pequeña fracción del alimento potencial obtenido de los autótrofos puede convertirse en producción animal, asumiendo un consumo del 100%. Con buenas estimaciones de la producción primaria y secundaria, y de la eficiencia ecológica, podemos calcular la fracción de la producción primaria que realmente consumen los pastores. Esta fracción no es necesariamente del 100% y puede proporcionar información considerable sobre los efectos descendentes de los animales en sus recursos alimenticios. y de las eficiencias ecológicas, podemos calcular la fracción de la producción primaria realmente consumida por los pastores. Esta fracción no es necesariamente del 100% y puede proporcionar información considerable sobre los efectos descendentes de los animales en sus recursos alimenticios. y de las eficiencias ecológicas, podemos calcular la fracción de la producción primaria realmente consumida por los pastores. Esta fracción no es necesariamente del 100% y puede proporcionar información considerable sobre los efectos descendentes de los animales en sus recursos alimenticios.
- ¿Es realista una eficiencia de producción bruta del 10% para todos los niveles tróficos? Parece que ocurre una gran variación, dependiendo de los tipos de organismos involucrados. Por ejemplo, mientras que P / A puede ser bastante alto, como el 40% para nuestros caracoles, P / A es comúnmente menor al 4% en animales endotérmicos (mamíferos, aves) lo que produciría un P / I mucho menor al 10%. Por otro lado, los depredadores invertebrados, como la libélula en la Tabla 1, típicamente tienen eficiencias de asimilación mucho más altas (70–90%) que los herbívoros (10–30%). Por lo tanto, es posible que> 30% de la producción de herbívoros se traduzca en producción de depredadores (p. Ej., 0,9 x 0,4 = 0,36). Bruce Wallace y sus colegas (1997) en realidad encontraron que la producción de invertebrados depredadores fue aproximadamente el 36% de la de los consumidores primarios en un ecosistema de arroyo durante muchos años. lo que implica que cerca del 100% de la producción del consumidor primario fue devorado por depredadores. En resumen, si bien podemos generalizar alrededor de un 10% de eficiencia de transferencia, en realidad es extremadamente variable (p. Ej., Humphreys 1979).
- ¿Cómo se relaciona la producción anual con la biomasa anual media? La respuesta a esta pregunta proporciona una estimación directa de las tasas de renovación de la biomasa y su relación con la duración de la vida. Los estudios han demostrado que si una población tiene una vida útil de un año, entonces la relación entre la producción anual y la biomasa media a lo largo del año (P / B) es aproximadamente 5 (Waters 1969). Además, la relación está inversamente relacionada con la esperanza de vida. Un período de vida de cinco años daría como resultado un P / B anual de aproximadamente 1. Un período de vida de treinta días daría como resultado un P / B anual de aproximadamente 60. Por lo tanto, puede ocurrir una gran variación en P / B anual que representa el Tasa de rotación de biomasa: un P / B anual de 30 significa que la población reemplaza su propia biomasa treinta veces en el transcurso de un año a través de los procesos de crecimiento y muerte.

Figura 3: Producción semanal de los tres principales taxones de libélulas durante un año en un estanque de Carolina del SurBasado en datos de Benke 1976.¿Cómo varía la producción en un año y qué nos puede decir esto? Aunque los estudios de producción típicos utilizan valores anuales, la Figura 3 muestra la producción semanal de la libélula Epithecade la Tabla 1 durante todo un año, así como para las otras dos libélulas más productivas del estanque. Obviamente, estas mediciones a corto plazo son más informativas que los valores anuales. Claramente, todas las especies tienen más producción durante el verano que en el invierno, un reflejo de mayores tasas de crecimiento a temperaturas más altas. El examen de tales patrones a veces sugiere una partición de nichos a lo largo de la dimensión temporal entre especies, pero eso no es evidente aquí. Sin embargo, dado que el consumo está directamente relacionado con la producción, estos patrones temporales sugieren una depredación mucho mayor por parte de las libélulas sobre sus presas (mosquitos quironómidos y efímeras) durante los meses de verano.- ¿Cuál es la importancia relativa de las especies potencialmente competidoras en una comunidad? La producción es una variable de respuesta particularmente útil para este propósito porque es un reflejo del éxito de una especie y su función (es decir, cuánta comida consume y cuánta comida puede contribuir potencialmente a sus depredadores). En el caso de las tres libélulas, estas especies son claramente depredadores co-dominantes en el estanque con producciones que van desde solo 1,93 a 2,27 gm -2 y -1. Se estimó que estas tres especies contribuían con alrededor del 75% de la producción total de libélulas en el estanque, a pesar de que había al menos otras diez especies de libélulas presentes. De manera similar, es posible comparar una población de una especie con otra de la misma especie en hábitats, comunidades o tratamientos experimentales.
- ¿Cómo se pueden cuantificar las redes alimentarias utilizando la producción? Las redes alimentarias basadas en especies son un tema de gran interés en la ecología, pero hay varias formas en las que se pueden construir. Bruce Wallace y yo desarrollamos un método para cuantificar los flujos de ingestión entre poblaciones utilizando una combinación de análisis de producción y de intestino (Benke y Wallace 1997). Al construir tales redes, es necesario hacer suposiciones de eficiencia ecológica a partir de la literatura científica (ver 1 y 2 arriba). La Figura 4 muestra una red de flujo cuantitativo parcial que se centra en las moscas caddis de un conjunto de insectos acuáticos en madera sumergida en un río. Tenga en cuenta que los flujos varían en más de 500 veces y que varias especies consumen cantidades significativas de presas animales. Claramente, estas moscas caddis no caen fácilmente en un solo nivel trófico.

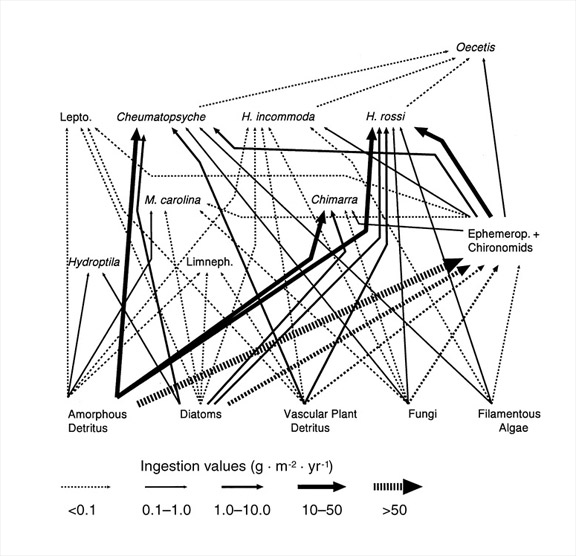
Figura 4: Red trófica cuantitativa para los principales taxones de caddisfly en el hábitat de madera sumergida en un ríoEl grosor de la línea indica la magnitud de los flujos de ingestión (g masa seca m -2 y -1 ). Las líneas discontinuas más gruesas son aproximaciones de flujos a efímeras (Ephemerop) y mosquitos (Chironomids). Lepto. son miembros de la familia Caddisfly Leptoceridae y Limneph. son miembros de la familia Limnephilidae. Todos los demás taxones son de género o especie. (Figura de Benke y Wallace 1997)
En resumen, la producción secundaria es una variable poblacional integral que incorpora densidad, biomasa, tasas de crecimiento individual, tasas de mortalidad de la población y esperanza de vida. No solo se ha utilizado en estudios de flujo de energía de los ecosistemas, sino que ha abordado una amplia variedad de cuestiones ecológicas. A menudo es superior a la densidad o la biomasa por sí solas como variable de respuesta porque es una medida de función más que de estructura.
