Fijación biológica de nitrógeno

El nitrógeno es un elemento limitante crítico para el crecimiento y la producción de las plantas. Es un componente principal de la clorofila, el pigmento más importante necesario para la fotosíntesis, así como los aminoácidos, los componentes clave de las proteínas. También se encuentra en otras biomoléculas importantes, como ATP y ácidos nucleicos. Si bien es uno de los elementos más abundantes (predominantemente en forma de gas nitrógeno (N 2) en la atmósfera de la Tierra), las plantas solo pueden utilizar formas reducidas de este elemento. Las plantas adquieren estas formas de nitrógeno «combinado» mediante: 1) la adición de amoníaco y / o fertilizante nitrato (del proceso Haber-Bosch) o estiércol al suelo, 2) la liberación de estos compuestos durante la descomposición de la materia orgánica, 3) la conversión de nitrógeno atmosférico en compuestos mediante procesos naturales, como rayos, y 4) fijación biológica de nitrógeno (Vance 2001).
La fijación biológica de nitrógeno (BNF), descubierta por Beijerinck en 1901 (Beijerinck 1901), es llevada a cabo por un grupo especializado de procariotas. Estos organismos utilizan la enzima nitrogenasa para catalizar la conversión de nitrógeno atmosférico (N 2 ) en amoniaco (NH 3 ). Las plantas pueden asimilar fácilmente el NH 3para producir las biomoléculas nitrogenadas antes mencionadas. Estos procariotas incluyen organismos acuáticos, como cianobacterias, bacterias del suelo de vida libre, como Azotobacter , bacterias que forman relaciones asociativas con plantas, como Azospirillum , y lo más importante, bacterias, como Rhizobium y Bradyrhizobium , que forman simbiosis con leguminosas y otras plantas (Postgate 1982).
El proceso de fijación del carbono
La reducción del nitrógeno atmosférico es un proceso complejo que requiere una gran cantidad de energía para continuar (Postgate 1982). La molécula de nitrógeno está compuesta por dos átomos de nitrógeno unidos por un triple enlace covalente, lo que hace que la molécula sea altamente inerte y no reactiva. La nitrogenasa cataliza la ruptura de este enlace y la adición de tres átomos de hidrógeno a cada átomo de nitrógeno.
Los microorganismos que fijan nitrógeno requieren 16 moles de trifosfato de adenosina (ATP) para reducir cada mol de nitrógeno (Hubbell y Kidder, 2009). Estos organismos obtienen esta energía oxidando moléculas orgánicas. Los microorganismos no fotosintéticos de vida libre deben obtener estas moléculas de otros organismos, mientras que los microorganismos fotosintéticos, como las cianobacterias, utilizan azúcares producidos por fotosíntesis. Los microorganismos asociativos y simbióticos fijadores de nitrógeno obtienen estos compuestos de las rizosferas de sus plantas hospedantes (National Research Council 1994, Hubbell & Kidder 2009).
Las industrias utilizan el proceso de Haber-Bosch para reducir el nitrógeno esencialmente de la misma manera. La agricultura convencional ha dependido de este proceso para producir el fertilizante comercial necesario para producir la mayoría de los cultivos híbridos del mundo. Pero este enfoque tiene muchas consecuencias, incluido el uso de combustibles fósiles para la energía necesaria para producir este fertilizante, las emisiones de dióxido de carbono y la contaminación resultantes de la quema de estos combustibles y los efectos adversos en la salud humana (Vitousek 1997).
El uso excesivo de estos fertilizantes químicos ha provocado una alteración del ciclo del nitrógeno y, en consecuencia, la contaminación de las aguas superficiales y subterráneas. El aumento de las cargas de fertilizantes nitrogenados en el agua dulce, así como en los ecosistemas marinos, ha provocado la eutrofización, el proceso por el cual estos sistemas tienen una proliferación de microorganismos, especialmente algas. Este «enverdecimiento» de la columna de agua ha provocado una disminución de los niveles de oxígeno disuelto (OD) en las aguas del fondo a medida que mueren las algas planctónicas y alimentan la respiración microbiana. Estos niveles reducidos de OD resultan en una mortalidad masiva de organismos acuáticos y crean las llamadas zonas muertas, áreas donde se puede encontrar poca o ninguna vida acuática (Figura 2). Desde la década de 1960, las zonas muertas han aumentado exponencialmente en todo el mundo y ahora se han documentado en más de 400 sistemas, afectando a más de 245, 000 kilómetros cuadrados de regiones costeras (Díaz & Rosenberg 2008, Figura 3). Este fenómeno ahora se considera el factor de estrés clave en los ecosistemas marinos.
Imagen en color verdadero de la deposición de sedimentos del río Mississippi en el Golfo de México
.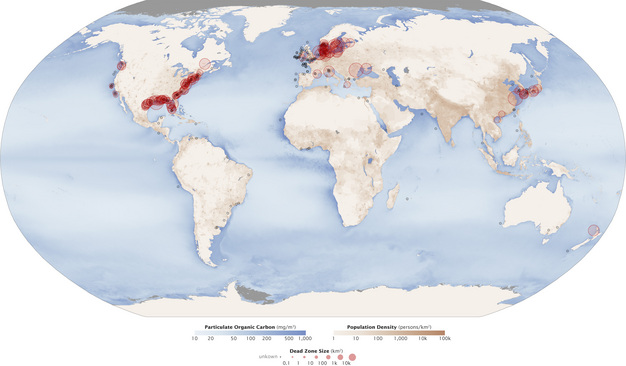
Zonas acuáticas muertas en todo el mundo.
Fijación de nitrógeno por heterótrofos de vida libre
Muchas bacterias heterótrofas viven en el suelo y fijan niveles significativos de nitrógeno sin la interacción directa con otros organismos. Ejemplos de este tipo de bacterias fijadoras de nitrógeno incluyen especies de Azotobacter , Bacillus , Clostridium y Klebsiella . Como se señaló anteriormente, estos organismos deben encontrar su propia fuente de energía, generalmente oxidando moléculas orgánicas liberadas por otros organismos o por descomposición. Hay algunos organismos de vida libre que tienen capacidades quimiolitotróficas y, por lo tanto, pueden utilizar compuestos inorgánicos como fuente de energía.
Debido a que la nitrogenasa puede ser inhibida por el oxígeno, los organismos de vida libre se comportan como anaerobios o microaerófilos mientras fijan nitrógeno. Debido a la escasez de fuentes de energía y carbono adecuadas para estos organismos, su contribución a las tasas globales de fijación de nitrógeno generalmente se considera menor. Sin embargo, un estudio reciente en Australia de un sistema de agricultura de rotación intensiva de trigo demostró que los microorganismos de vida libre contribuían con 20 kilogramos por hectárea por año a las necesidades de nitrógeno a largo plazo de este sistema de cultivo (30-50% de las necesidades totales; Vadakattu & Paterson 2006). El mantenimiento del rastrojo de trigo y la labranza reducida en este sistema proporcionó el ambiente con alto contenido de carbono y bajo nitrógeno necesario para optimizar la actividad de los organismos de vida libre.
Fijación asociativa de nitrógeno
Las especies de Azospirillum pueden formar asociaciones cercanas con varios miembros de las Poaceae (pastos), incluidos cultivos de cereales de importancia agronómica, como arroz, trigo, maíz, avena y cebada. Estas bacterias fijan cantidades apreciables de nitrógeno dentro de la rizosfera de las plantas hospedantes. Se ha informado de eficiencias de 52 mg de N 2 g -1 de malato (Stephan et al . 1979). El nivel de fijación de nitrógeno está determinado por varios factores, incluida la temperatura del suelo ( Azospirillum especies prosperan en ambientes más templados y / o tropicales), la capacidad de la planta huésped para proporcionar un ambiente rizosférico con baja presión de oxígeno, la disponibilidad de fotosintetos del huésped para las bacterias, la competitividad de las bacterias y la eficiencia de la nitrogenasa (Vlassak Y Reynders, 1979).
Fijación simbiótica de nitrógeno
Muchos microorganismos fijan el nitrógeno de forma simbiótica al asociarse con una planta huésped. La planta proporciona azúcares de la fotosíntesis que son utilizados por el microorganismo fijador de nitrógeno para obtener la energía que necesita para la fijación de nitrógeno. A cambio de estas fuentes de carbono, el microbio proporciona nitrógeno fijo a la planta huésped para su crecimiento.
Un ejemplo de este tipo de fijación de nitrógeno es la simbiosis del helecho acuático Azolla con una cianobacteria Anabaena azollae . Anabaena coloniza las cavidades formadas en la base de Azollafrondas. Allí, las cianobacterias fijan cantidades significativas de nitrógeno en células especializadas llamadas heterocistos. Esta simbiosis se ha utilizado durante al menos 1000 años como biofertilizante en arrozales de humedales en el sudeste asiático. Los arrozales suelen estar cubiertos de «floraciones» de Azolla que fijan hasta 600 kg de N ha -1 año -1 durante la temporada de crecimiento (Postgate 1982, Fattah 2005).
Otro ejemplo es la simbiosis entre árboles y arbustos actinorriza, como el aliso ( Alnus sp.), Con el actinomiceto Frankia. Estas plantas son nativas de América del Norte y tienden a prosperar en ambientes pobres en nitrógeno. En muchas áreas son los fijadores de nitrógeno no leguminosos más comunes y, a menudo, son las especies pioneras en las comunidades de plantas sucesivas. Las plantas actinorrizas se encuentran en muchos ecosistemas, incluidos los ambientes alpino, xérico, chapparal, forestal, glacial, ribereño, de dunas costeras y tundra ártica (Benson y Silvester, 1993).
Aunque los socios simbióticos descritos anteriormente juegan un papel importante en la ecología mundial de la fijación de nitrógeno, las asociaciones simbióticas de fijación de nitrógeno más importantes son, con mucho, las relaciones entre leguminosas y Rhizobium y Bradyrhizobium.bacterias. Las leguminosas importantes que se utilizan en los sistemas agrícolas incluyen alfalfa, frijoles, trébol, caupí, altramuces, maní, soja y arvejas. De las leguminosas en la producción agrícola, la soja se cultiva en el 50% del área mundial dedicada a las leguminosas y representa el 68% de la producción mundial total de leguminosas (Vance 2001).
Formación de nódulos de leguminosas
Las bacterias Rhizobium o Bradyrhizobium colonizan el sistema de raíces de la planta huésped y hacen que las raíces formen nódulos para albergar las bacterias (Figura 4). Luego, las bacterias comienzan a fijar el nitrógeno requerido por la planta. El acceso al nitrógeno fijo permite que la planta produzca hojas fortificadas con nitrógeno que se pueden reciclar en toda la planta. Esto permite que la planta aumente la capacidad fotosintética, lo que a su vez produce semillas ricas en nitrógeno. Las consecuencias de que las leguminosas no se nodulen pueden ser bastante dramáticas, especialmente cuando las plantas crecen en suelos pobres en nitrógeno. Las plantas resultantes son típicamente cloróticas, con bajo contenido de nitrógeno y producen muy pocas semillas (Figuras 5 y 6).

Nodulación extensa de una raíz de maní después de la inoculación con la cepa 32H1 de Bradyrhizobium .
Soja mutante no nodulada (primer plano) con soja normal, nodulada (fondo).
Comparación de plantas de maní con y sin Bradyrhizobia. Las plantas están (de izquierda a derecha), no inoculadas con Bradyrhizobium , inoculadas con Bradyrhibium , maní mutante no nodulante inoculado con Bradyrhizobium y maní mutante no nodulante sin inocular con Bradyrhizobium .
El proceso de nodulación ilustra una interacción orquestada entre la bacteria y la planta huésped (Napoli & Hubbell 1975, Kamst et al . Van Rhyn & Vanderleyden 1995, Cheng & Walker 1998). El proceso comienza cuando los rizobios son atraídos por los flavonoides liberados por las raíces de la leguminosa huésped. Para las legumbres como la alfalfa, el trébol y la soja (otras como los altramuces y los cacahuetes forman nódulos de otras formas), las bacterias comienzan a adherirse a las extensiones de las células epidérmicas de la raíz llamadas pelos de la raíz. El proceso de unión es en realidad un proceso de dos pasos en el que las bacterias se adhieren primero usando un Ca 2+– proteína de unión llamada ricadhesina. Después de que las bacterias se acumulan y se anclan a la superficie del pelo de la raíz, se crea una unión más firme que involucra lectinas y / o firbrils y fimbrias de celulosa producidas por la planta huésped y las bacterias, respectivamente.
Luego, la leguminosa huésped detecta las sustancias químicas producidas por los rizobios llamados factores Nod que hacen que los pelos de la raíz colonizados se ricen y formen lo que se llama un cayado de pastor. Luego, los rizobios penetran los pelos de la raíz y generalmente forman una estructura tubular llamada hilo de infección. Una vez que las bacterias llegan a la raíz, estimulan las divisiones de las células corticales que conducen a la formación de un nódulo. A medida que el nódulo comienza a formarse, las bacterias quedan rodeadas por una membrana de origen vegetal y se liberan dentro de las células vegetales que forman el nódulo. Posteriormente, las bacterias pierden sus paredes celulares y experimentan un cambio profundo en la morfología celular para formar células ramificadas grandes de forma irregular llamadas bacteroides. Entonces dependen por completo de la planta huésped para sus necesidades energéticas. A cambio, las bacterias fijan nitrógeno para la planta.
La interacción entre las bacterias y la leguminosa huésped es tan intrincada que un Rhizobium o Bradyrhizobium en particular solo nodulará un número selecto de géneros de plantas. Por ejemplo, Rhizobium melilotii solo nodulará alfalfa, mientras que Rhizobium leguminosarum biovar trifolii solo nodulará trébol ( Trifolium ). Esta especificidad del huésped se refiere a la señalización celular del grupo de inoculación cruzada entre la bacteria y la leguminosa huésped. Los factores Nod antes mencionados se han identificado como oligosacáridos de lipoquición. Las variaciones en las estructuras de estos oligosacáridos determinan la especificidad del hospedador para la bacteria. Los principales grupos de inoculación cruzada se enumeran en la Tabla 1.
| Cosecha | Bacterias noductoras |
| Alfalfa | Sinorhizobium melilotii |
| Frijoles | Rhizobium legumninosarum biovar phaseoli y Rhizobium tropici |
| Trébol | Rhizobium leguminosarum biovar trifolii |
| Loto | Mesorhizobium loti |
| Chícharos | Rhizobium leguminosarum biovar viceae |
| Haba de soja | Bradyrhizobium japonicum , Bradyrhizobium elkanii , Rhizobium fredii |
| Sesbania | Azorhizobium caulinodans |
| Cuadro 1. Principales grupos de inoculación cruzada de leguminosas. |
Otro ejemplo de la intrincada relación entre los rizobios y la leguminosa huésped es la producción de leghemoglobina (Appleby 1984). Esta molécula es similar en estructura y función a la hemoglobina que se encuentra en la sangre humana. Solo se produce en los nódulos radiculares en pleno funcionamiento y funciona para unir y regular los niveles de oxígeno en el nódulo. Debido a que la enzima nitrogenasa es sensible al oxígeno, el oxígeno libre en el citoplasma de las células nódulos inhibiría los niveles de fijación de nitrógeno en el nódulo. La leghemoglobina parece transportar suficiente oxígeno para permitir que los rizobios realicen la respiración celular, pero no demasiado para inhibir la acción de la nitrogenasa. Esta proteína hemo es producida conjuntamente por la leguminosa y la bacteria; la leguminosa produce la apoproteína mientras que la bacteria produce el hemo (anillo de porfirina unido a un átomo de hierro).
Resumen
El nitrógeno es un nutriente esencial para el crecimiento y desarrollo de las plantas, pero no está disponible en su forma más común como nitrógeno atmosférico. Las plantas, en cambio, dependen de formas combinadas o fijas de nitrógeno, como el amoníaco y el nitrato. Gran parte de este nitrógeno se proporciona a los sistemas de cultivo en forma de fertilizantes nitrogenados producidos industrialmente. El uso de estos fertilizantes ha provocado problemas ecológicos en todo el mundo, como la formación de zonas muertas costeras. La fijación biológica de nitrógeno, por otro lado, ofrece un medio natural de proporcionar nitrógeno a las plantas. Es un componente crítico de muchos ecosistemas acuáticos y terrestres en nuestra biosfera.
